An Outflow Tract Myocardium-Specific Enhancer at the Sema3c Locus During Heart Development.
Sema3c is specifically expressed in the cardiac outflow tract (OFT) of the developing mouse heart and has been implicated in OFT polarization and great artery formation. However, the regulatory basis underlying its spatially restricted expression remains unclear. To investigate the mechanisms underlying OFT-specific Sema3c expression, we utilized chromatin accessibility data from distinct segments of the developing heart and identified a differentially accessible region as an OFT-specific Sema3c enhancer candidate. Unlike previously characterized Sema3c enhancers, this region is located distal to the transcription start site. Reporter analysis using transgenic mouse embryos demonstrated that this region exhibits transcriptional activity from E8.5 onward and remains specifically active in the OFT myocardium throughout heart development. We further defined a minimal 603 bp enhancer whose activity depends on GATA binding sites. This enhancer provides insight into the mechanisms underlying spatially restricted Sema3c expression involved in OFT development.
Introduction
Congenital heart disease (CHD) affects approximately 0.8% of all live births (Mitchell et al.1971; Reller et al.2008; Van Der Linde et al.2011), among which defects involving the cardiac outflow tract (OFT) and the great arteries account for an estimated 30% of cases (Egbe et al.2014; Thom et al.2006). During mammalian heart development, the OFT plays a critical role in connecting the developing ventricles to the great arteries and is essential to establish early blood circulation (Neeb et al.2013; Garry and Olson2006). The cellular components of the OFT are primarily derived from the anterior second heart field (SHF) and neural crest cells (NCCs) (Kelly et al.2014; Meilhac and Buckingham2018). At the four‐chamber stage of heart development, polarization within the OFT and subsequent septation give rise to the separation of the aorta and pulmonary trunk (Hikspoors et al.2022; Plein et al.2015).Sema3c, a member of the class 3 semaphorin family (Valdembri et al.2016), is specifically expressed in the OFT during heart development, and loss ofSema3cin mice results in abnormal formation of aorta and pulmonary trunk seen in CHDs including persistent truncus arteriosus (PTA) (Feiner et al.2001). Thus, considerable efforts have been devoted to elucidating the transcriptional regulation ofSema3cexpression. Loss‐of‐function experiments revealed thatTbx1 and Nkx2‐5are involved in OFT‐specific expression ofSema3c(Théveniau‐Ruissy et al.2008; Rana et al.2014; Yamaguchi et al.2023). In addition, two enhancers regulatingSema3chave been reported so far: an intronic enhancer responsive to GATA6 (Lepore et al.2006; Kodo et al.2009) and a proximal enhancer regulated by Foxc1/2 (Kodo et al.2017). However, mechanisms explaining OFT‐specificSema3cexpression remain poorly understood.
We have been conducting a genome‐wide search for enhancers that regulate gene expression in developing hearts. While the details will be presented in another manuscript (Harada et al. in preparation), the present study aims to examine an OFT‐specific differentially accessible region (DAR) located near theSema3clocus as an enhancer candidate. UsinglacZreporter transgenic mouse lines, we demonstrated that 1842 bp OFT‐specific DAR exhibits enhancer activity within the OFT myocardium throughout cardiac development. We further defined a minimal 603 bp region that retains OFT‐specific enhancer activity and demonstrated that its activity depends on GATA binding sites but not FOX binding sites. Together, our findings identify an OFT myocardium‐specific enhancer associated with theSema3clocus and provide a clue for investigating the unique transcriptional regulatory programs that underlie OFT development.
Results
Identification of anOFT‐Specific Accessible Chromatin Region as a NovelSema3cEnhancer Candidate
To investigate how OFT‐specific expression ofSema3cis regulated during heart development, we focused on OFT‐specific DAR detected at the looping stage in mice (E9.5). Comparing chromatin accessibility in the genomes of four anatomical segments: OFT, ventricle (V), atrioventricular canal (AVC), and atrium (A), we found the DAR as aSema3cenhancer candidate. This region spans 1842 bp and is located approximately 15 kb upstream of theSema3ctranscription start site (Figure1A).Sema3cis the closest gene to this enhancer candidate, and the other neighbor genes have not been reported to be expressed in the developing heart. Comparative genomics analysis using the Vertebrate Multiz Alignment and Conservation tracks of the UCSC Genome Browser revealed that this enhancer candidate is highly conserved among mammals, including humans, but is not conserved in non‐mammalian vertebrates (Figure1B).
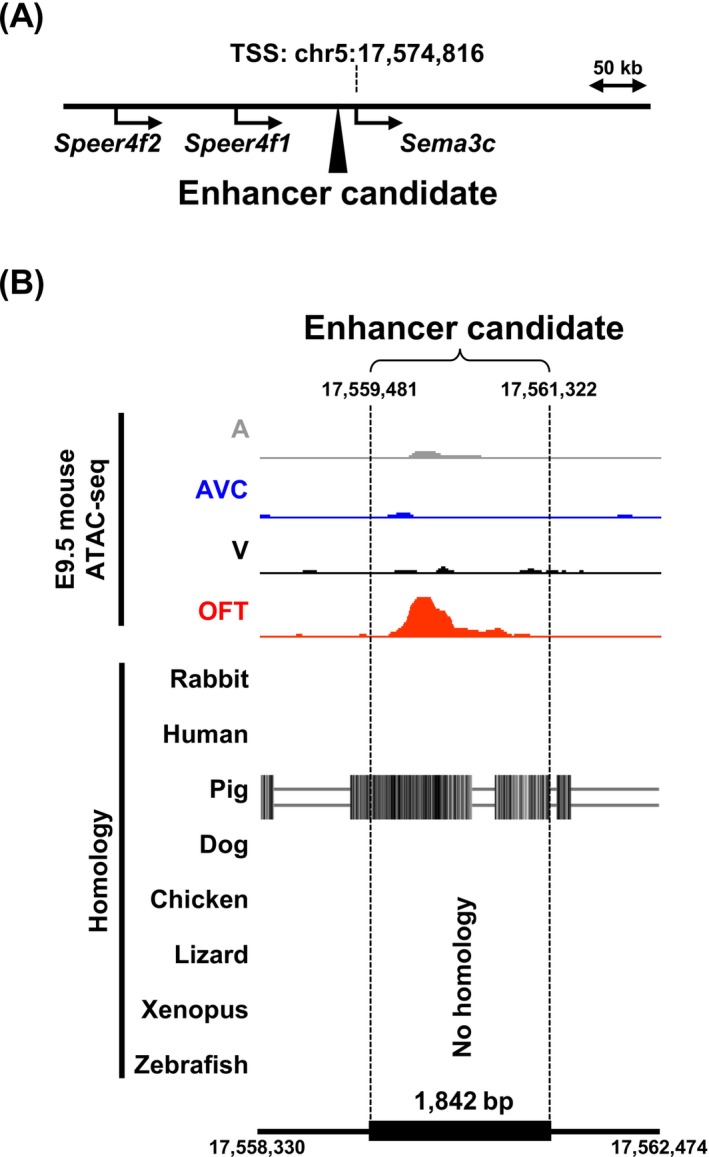
Identification of aSema3cenhancer candidate from an OFT‐specific accessible chromatin region. (A) The closest OFT‐specific DAR upstream ofSema3cwas identified as an enhancer candidate. The mouse genome assembly version, GRCm38/mm10 was used. (B) ATAC‐seq data of the 1842 bp enhancer candidate located at chr5:17,559,481–17,561,322 are compared among four independent anatomical segments of the E9.5 mouse heart: Atrium (A), atrioventricular canal (AVC), ventricle (V), and outflow tract (OFT). Comparative genomics analysis using the UCSC Genome Browser showed that this candidate exhibits mammalian‐conserved sequence homology.
To assess whether this enhancer candidate possesses transcriptional activity,lacZreporter transgenes were generated by pronuclear injection, resulting in three independent transgenic mouse lines (Line 1, Line 2, and Line 3). X‐gal staining performed at 37°C for 2 h on E9.5 embryos from all three lines revealed reproducible reporter activity specifically in the OFT of looping‐stage hearts (Figure2A). Further analysis of cryosections prepared from X‐gal stained E9.5 embryos demonstrated that reporter activity driven by the enhancer candidate was localized to the myocardial layer of the OFT (Figure2B). Together, these results indicate that the newly identified OFT‐specific DAR represents an enhancer that may contribute to the regulation ofSema3cexpression in the OFT myocardium during heart development.
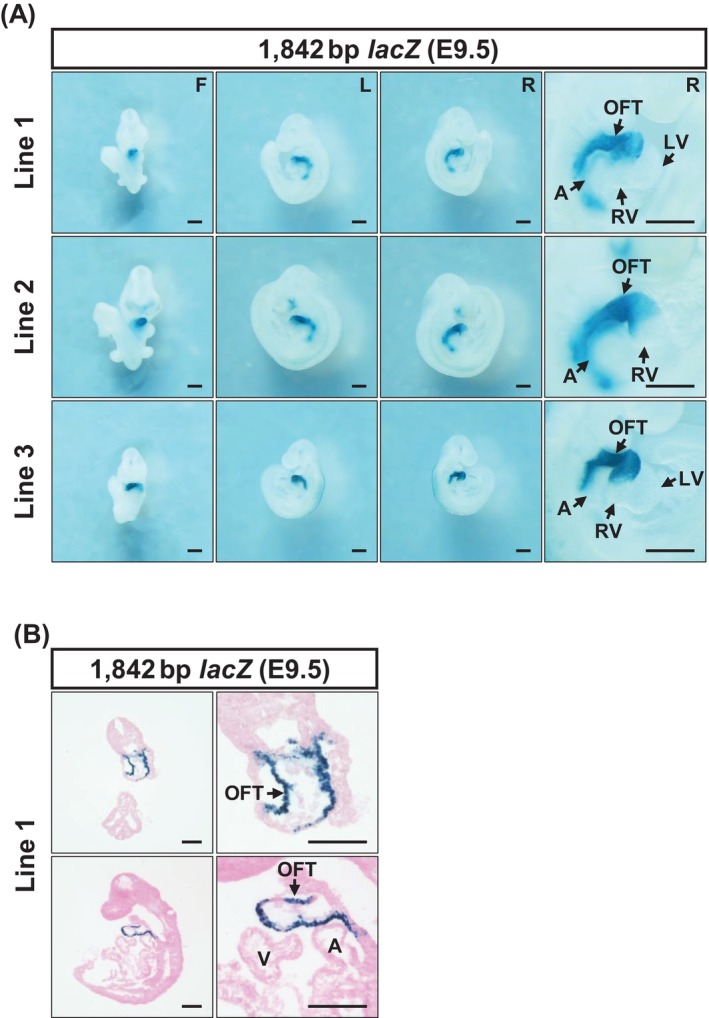
The 1842 bp enhancer candidate exhibits OFT‐specific transcriptional activity in the E9.5 mouse myocardium. (A) Three independent lines of 1842 bp enhancer‐hsp68‐lacZtransgenic reporter mouse were analyzed. Representative images of whole mount X‐gal staining (37°C°C for 2 h) are shown. (B) To further assess the cellular localization of reporter activity observed in OFT, cryosections were prepared.LacZactivity driven by the 1842 bp DAR was localized to the myocardial layer of the OFT. A, atrium; F, front view; L, left view; LV, left ventricle; OFT, outflow tract; R, right view; RV, right ventricle. Scale bar: 500 μm.
The Enhancer Candidate Exhibits Sustained Myocardium‐Specific Activity DuringOFTDevelopment
To characterize the temporal dynamics of enhancer activity across different stages of OFT development,lacZreporter activity was examined from E7.5 to E12.5 at 24 h intervals, using one of the transgenic mouse lines (Line1). No reporter activity was detected in embryos at E7.5 stage (FigureS1). At the early stage of OFT formation (E8.5), X‐gal staining performed at 37°C for 24 h revealed reporter activity in a segment corresponding to the early OFT (Figure3A). Compared with later stages, reporter activity at this stage appeared relatively weak, and no reporter activity was detected in other cardiac segments nor outside the heart at this stage (Figure3A).

The 1842 bp enhancer exhibits temporally sustained and OFT‐specific transcriptional activity during heart development. (A) Representative images of whole‐mount X‐gal staining of 1842 bp enhancer‐hsp68‐lacZtransgenic embryos are shown. At E8.5, reporter activity was detected in the early OFT of the developing heart tube. During the looping stages (E9.5–E11.5), enhancer activity was observed in the OFT but not in the RV. Reporter activity was also detected in a domain corresponding to the SHF at E9.5 and E10.5 (arrowheads). At the four‐chamber stage (E12.5), enhancer activity remained localized to the OFT. Although weak reporter activity was observed in the right atrium in a subset of embryos, analysis of additional independent transgenic lines confirmed that enhancer activity was not reproducibly detected in the right atrium. The E9.5 image is the same as shown in Figure2A. X‐gal staining was performed at 37°C°C for 2 h (E9.5) or 24 h (E8.5 and E10.5–E12.5). (B) Cryosections prepared at E10.5–E11.5 exhibited that reporter activity was localized to the myocardial layer of the OFT. A, atrium; F, front view; L, left view; LV, left ventricle; OFT, outflow tract; R, right view; RA, right atrium; RV, right ventricle; V, ventricle. Scale bar: 500 μm.
During the OFT elongation stages (E9.5–E11.5), X‐gal staining (E9.5, 37°C for 2 h; E10.5 and E11.5, 37°C for 24 h) revealed highly consistent and spatially restricted enhancer activity (Figure3A). At these stages, reporter activity was detected exclusively in the OFT and was absent from the right ventricle, which is also derived from the SHF. Outside the heart, enhancer activity was also observed at E9.5 and E10.5 around a posterior wall region corresponding to the dorsal pericardium (arrowheads), a structure closely associated with OFT and right ventricular development (Figures2Aand3A) (Stefanovic et al.2020).
At E12.5, corresponding to the early four‐chamber stage, X‐gal staining (37°C for 24 h) demonstrated that enhancer activity remained restricted to the OFT (Figure3A). No reproducible signal was observed in the right ventricle, consistent with the spatial pattern observed at earlier looping stages. Although embryos from Line 1 exhibited weak reporter activity in the right atrium at E12.5, analysis of Line 2 confirmed that enhancer activity was not reproducibly detected in the right atrium at this stage (FigureS2).
Cryosection analysis further demonstrated that, from E9.5 to E12.5, enhancer‐driven reporter activity was confined to the myocardial layer of the OFT and did not extend into the right ventricular myocardium (Figures2Band3B). Notably, reporter activity was not detected in the OFT cushions, despite endogenousSema3cexpression in this region (Figures2Band3B). Considering these results, we can conclude that the enhancer candidate serves as theSema3cOFT enhancer exhibiting sustained OFT myocardium‐specific activity.
A 603 bp Minimal Enhancer RetainsOFT‐Specific Transcriptional Activity
Although the 1842 bp enhancer exhibited clear transcriptional activity, its length is relatively long compared with typical functional enhancers (Blackwood and Kadonaga1998; Li and Wunderlich2017), prompting us to investigate whether a shorter fragment could retain enhancer activity. ATAC‐seq peak profile of the 1842 bp enhancer region showed that chromatin accessibility was not uniform across the region, displaying an asymmetry with greater accessibility in its distal half (Figure4A). To test whether a shorter enhancer fragment could retain activity, we first removed the low‐accessibility region of the 1842 bp enhancer and generated an 881 bp enhancer candidate fragment. F0 reporter analysis at E9.5, with X‐gal staining (37°C for 24 h), confirmed that this 881 bp fragment exhibited OFT‐specific activity comparable to that of the 1842 bp enhancer (Figure4). The result above supports our hypothesis that enhancer activity correlates with chromatin accessibility.
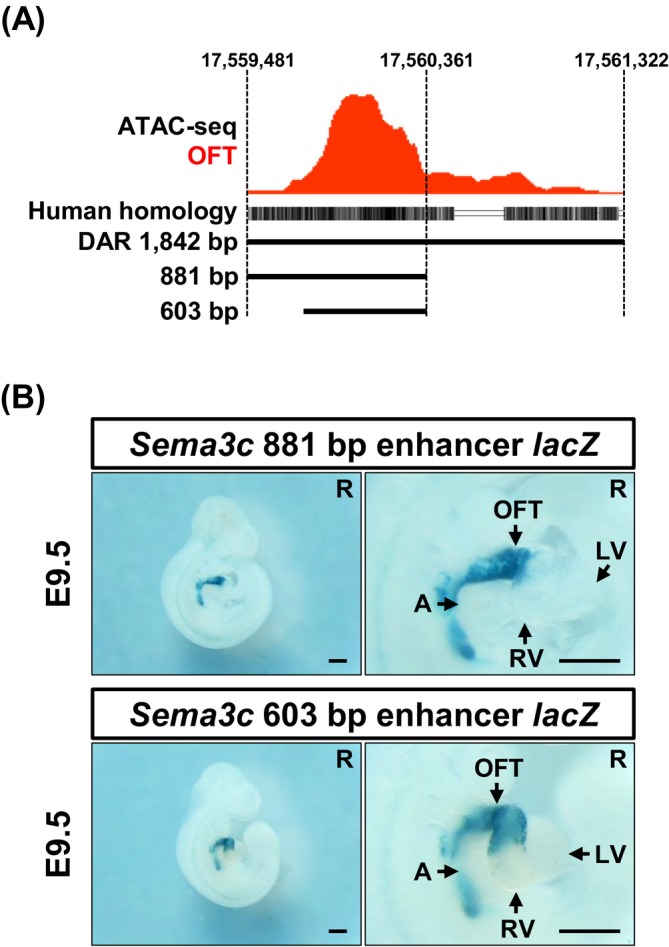
A 603 bp minimal enhancer exhibits OFT‐specific activity. (A) Two truncated enhancer fragments (881 and 603 bp) are derived from the OFT‐specific DAR based on chromatin accessibility profiles. (B) Both minimal enhancer fragments drove OFT‐specific reporter activity in F0 transient transgenic embryos. The spatial pattern of reporter activity driven by the 881 bp (n= 2/2) and 603 bp (n= 5/8) fragments is shown. A, atrium; LV, left ventricle; OFT, outflow tract; R, right view; RV, right ventricle. Scale bar: 500 μm.
To define the minimal region required for enhancer activity, we further removed the low‐accessibility portion and generated a 603 bp enhancer candidate fragment (Figure4A). F0 reporter analysis with X‐gal staining (37°C for 6 h) revealed that this 603 bp fragment exhibited an OFT‐specificlacZexpression pattern (E9.5) equivalent to those of the 1842 and 881 bp enhancers (Figure4). Further deletion attempts failed to produce this pattern comparable to that of the 1842 bp enhancer (FigureS3). These findings indicate that the 603 bp fragment represents the minimalSema3cenhancer capable of driving OFT‐specific transcriptional activity.
GATABinding Site Dependent Activity of theSema3c603 bp Enhancer
Two cardiac enhancers regulatingSema3cexpression have been previously characterized: a GATA6‐dependent intronic enhancer (Lepore et al.2006; Kodo et al.2009) and a Foxc1/2‐responsive proximal enhancer (Kodo et al.2017). In contrast to these enhancers, both of which are located close to the transcription start site (TSS), the enhancer identified in this study is located at a distal position upstream of theSema3clocus (Figure5A). We next performed motif analysis of the distal 603 bpSema3cenhancer. Using the JASPAR 2024 database, we examined predicted binding motifs for FOX and GATA transcription factor families, which have been reported to regulate previously characterizedSema3cenhancers. Two GATA and one FOX binding motifs were identified within the 603 bp enhancer (Figure5B).
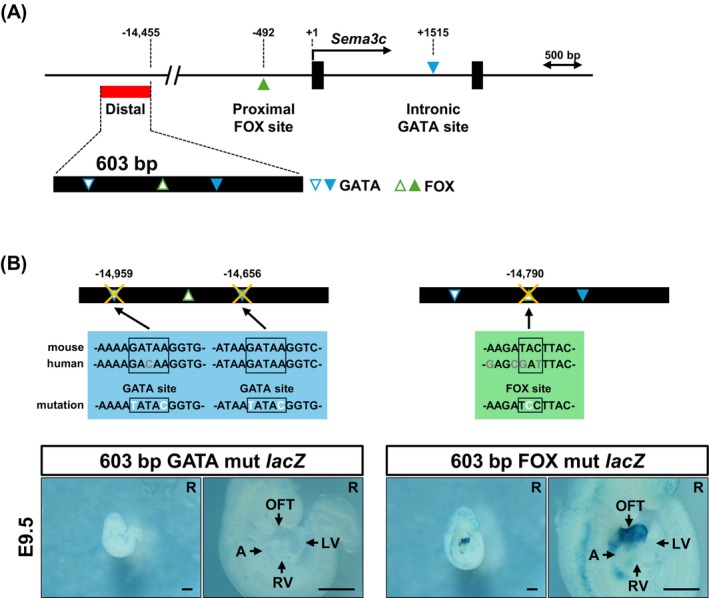
The activity of theSema3c603 bp enhancer depends on GATA transcription factor binding sites. (A) Genomic location of a novel distal enhancer ofSema3c. According to the previous reports, transcription factor binding sites were annotated in the 5′ promoter‐proximal region and within intron 1 of the mouseSema3cgene. The 603 bp enhancer (red box) identified in this study is located approximately 15 kb upstream of theSema3ctranscription start site. Motif analysis of the 603 bp enhancer using the JASPAR database (a relative profile score threshold of > 95%) identified enriched transcription factor binding motifs of GATA and FOX families. Open symbols indicate motifs specific to the mouse enhancer sequence, whereas filled symbols denote motifs conserved in humans. (B) The 603 bp enhancer contains two GATA binding sites. Mutation of these sites in F0 embryos resulted in a complete loss of enhancer activity. In contrast, mutation of a single FOX binding site in this enhancer did not affect reporter activity. X‐gal staining was performed at 37°C°C for 24 h (GATA mutation) and 6 h (FOX mutation). A, atrium; LV, left ventricle; OFT, outflow tract; R, right view. Scale bar: 500 μm.
Considering previous reports implicating GATA and FOX family transcription factors as upstream regulators ofSema3cenhancers (Lepore et al.2006; Kodo et al.2009,2017), we next examined the functional contribution of these motifs by introducing site‐directed mutations into the predicted binding sites within the 603 bp enhancer (Figure5B). Reporter constructs carrying mutations in either the GATA binding sites or the FOX site were analyzed by F0 transgenic reporter assays. Mutation of the two predicted GATA binding sites resulted in a complete loss of reporter activity in E9.5 embryos following X‐gal staining at 37°C for 24 h. This result indicated that the transcriptional activity of the 603 bp enhancer depends on intact GATA binding sites (Figure5B). In contrast, mutation of the single predicted FOX binding site did not abolish enhancer activity. In addition, no ectopiclacZexpression was observed in F0 reporter embryos carrying each mutant enhancer, compared to the wild‐type enhancer (Figure5B). Together, these results indicate that GATA binding sites are required for the transcriptional activity of the distalSema3c603 bp enhancer in the OFT myocardium.
Discussion
In this study, we validated an OFT‐specific DAR upstream of theSema3clocus (Figure1). Using transgeniclacZreporter mouse lines, we demonstrated that this region exhibits transcriptional activity in the cardiac OFT, thereby defining an OFT myocardium‐specific enhancer ofSema3c(Figures2and3). Through progressive truncation, we further defined a minimal 603 bp enhancer that retains OFT‐specific transcriptional activity (Figures4andS3). Mutation of GATA family transcription factor binding sites within this minimal region resulted in a complete loss of enhancer activity, indicating that enhancer function is dependent on GATA transcription factor binding (Figure5).
Temporal analysis of reporter embryos from E8.5 to E12.5 revealed that enhancer activity is initiated at the onset of OFT formation and subsequently maintained throughout its development, with activity consistently restricted to the OFT myocardium (Figure3). Although weak X‐gal signals were detected outside the OFT (Figure3), strong transcriptional activity within the heart was reproducibly confined to the OFT across independent transgenic lines (FigureS2). In future studies, deleting theSema3cOFT enhancer will be important to examine the changes inSema3cexpression and determine its potential impact on OFT morphogenesis. Notably, enhancer activity was not detected in the OFT cushions during the E9.5–E12.5 period (Figures2Band3B), despite endogenousSema3cexpression in this tissue (Plein et al.2015). This finding suggests thatSema3cexpression in the OFT myocardium and cushions is governed by distinct transcriptional regulatory mechanisms. Previous studies reported thatSema3cexpression in neural crest‐derived, Wnt1‐positive OFT cushions contributes to OFT septation (Plein et al.2015). Meanwhile,Sema3cis expressed in the SHF independently of Wnt signaling (Rammah et al.2022). Analysis of early developmental stages further showed that the 1842 bpSema3cenhancer is not active in the SHF outside the OFT at E8.5, but becomes detectable by E9.5 and persists through E10.5 (Figure3) (Kelly2023). Although the anterior SHF contributes to both the OFT and right ventricle (Meilhac and Buckingham2018; Verzi et al.2005; Stefanovic et al.2020), the 1842 bpSema3cenhancer did not exhibit activity in the right ventricle at any stage examined (Figure3). These observations suggest that cardiac progenitors in the anterior SHF may undergo further regional or lineage diversification during cardiac looping stages. Lineage‐tracing approaches may help to clarify the distinct developmental trajectories underlying OFT and right ventricular formation.
GATA6 and Foxc1/2 have been shown to regulateSema3ctranscription through previously characterized enhancer elements located near the transcription start site (Lepore et al.2006; Kodo et al.2009,2017). Our results show that the activity of the distal 603 bpSema3cOFT enhancer depends on GATA transcription factor binding sites (Figure5). Previous studies have demonstrated that GATA family transcription factors are closely associated with chromatin accessibility, and that loss of GATA factors leads to reduced or abolished accessibility at their target regulatory regions (Heslop et al.2021; Yamaguchi et al.2023). It is therefore possible that GATA transcription factors contribute to the establishment or maintenance of OFT‐specific chromatin accessibility at the distalSema3cenhancer. However, the presence of GATA binding sites alone cannot explain the OFT myocardium‐specific activity of the enhancer, as GATA factors are broadly involved in cardiac gene regulation (Charron and Nemer1999). In addition, weak and discontinuouslacZexpression was observed in several enhancer candidates shorter than 603 bp (FigureS3), although GATA transcription factor binding sites were present in these candidates. We therefore speculate that additional, yet unidentified regulatory factors cooperate with GATA transcription factors to confer the spatial specificity of this enhancer. Identifying such factors will be an important direction for future investigation. In contrast to the proximal enhancer reported by Kodo et al. (2017), mutation of the FOX site did not affect the activity of the 603 bpSema3cOFT enhancer (Figure5B). This may result from the lack of functionality at this FOX site, which is not conserved in humans, unlike the 603 bp enhancer exhibiting high sequence conservation across mammals. Furthermore, sinceFoxc1/2are expressed in the non‐myocardium of the OFT, they are unlikely to regulate this newly identifiedSema3cOFT enhancer, which is not active in the cushion. Previous studies have reported thatTbx1andNkx2‐5are associated withSema3cexpression in the OFT (Théveniau‐Ruissy et al.2008; Rana et al.2014; Yamaguchi et al.2023). These transcription factors, along with their downstream effectors, may cooperate with GATA factors to establish the OFT‐myocardium‐specific activity of this enhancer. Interestingly,Sema3cexpression is controlled by multiple enhancers with distinct regulatory upstream factors during OFT development.
Although the precise transcriptional regulatory mechanisms remain to be fully defined, the distinctive properties of theSema3cOFT enhancer provide a useful tool for dissecting cellular heterogeneity between OFT myocardium and OFT cushions, as well as between OFT myocardium and right ventricular myocardium. In addition, its activity within the SHF may facilitate further refinement of myocardial progenitor classification and lineage tracing in this region. Collectively, the enhancer identified in this study expands the current understanding of transcriptional regulation during OFT development.
Experimental Procedures
lacZReporter Analysis
Animal experiments were performed with the approval of the institutional animal care and use committee of Ritsumeikan University (BKC2023‐033). To generatelacZreporter transgenic (Tg) mice, linearized plasmid constructs were injected into B6D2F1/Slc (from Japan SLC Inc.) fertilized eggs.
Enhancer candidate–hsp68 promoter‐nls‐lacZtransgenic mice were maintained on a C57BL/6NCrSlc background (Japan SLC Inc.). Embryos were collected at the indicated developmental stages and genotyped as previously described.lacZreporter activity in embryonic hearts was detected by X‐gal staining (Ihara et al.2020).
For X‐gal staining, embryos at E8.5–E9.5 were pre‐fixed in 4% paraformaldehyde (PFA) in phosphate‐buffered saline (PBS) on ice for 15 min, whereas embryos at E10.5–E12.5 were pre‐fixed in 4% PFA/PBS on ice for 30 min. Following X‐gal staining, all embryos were post‐fixed in 4% PFA/PBS at 4°C for 16 h prior to imaging.
Plasmid Construction
Genomic DNA fragments corresponding to enhancer candidate regions for mouseSema3cwere amplified by PCR and cloned into the pT2AL200R175 plasmid containing the hsp68 promoter‐nls‐lacZreporter cassette using the In‐Fusion HD EcoDry Cloning Kit (Takara Bio, 639690).
Primer sequences used for PCR amplification (excluding In‐Fusion adapter sequences) were as follows:
for the 1842 bp enhancer, forward 5′‐GCTGACTGTGGGAGTCTCAA‐3′ and reverse 5′‐TTGCGGGAAAGACAAGGTCA‐3′;
for the 881 bp enhancer, forward 5′‐GCTGACTGTGGGAGTCTCAA‐3′ and reverse 5′‐CATTTAATAAGTTAACAAGA‐3′;
for the 603 bp enhancer, forward 5′‐CATACAGAAATGAAGATGGG‐3′ and reverse 5′‐CATTTAATAAGTTAACAAGA‐3′;
for the 603 bp enhancer GATA binding site mutation, pair 1 for first site, forward 5′‐CTTCCAGTAGAAAAATATACGGTGGCCCACACCCCCAGTG‐3′ and reverse 5′‐GGGGTGTGGGCCACCGTATATTTTTCTACTGGAAGTATTTATGG‐3′; pair 2 for second site, forward 5′‐TTCACATACAAATAATATACGGTCAGGGTCAGGATCCAC‐3′ and reverse 5′‐ATCCTGACCCTGACCGTATATTATTTGTATGTGAATTTCC‐3′;
for the 603 bp enhancer GATA binding site mutation, forward 5′‐CGTTTGACCTAAAGATCCTTACAAGTATGAGTTTATCATATG‐3′ and reverse 5′‐AACTCATACTTGTAAGGATCTTTAGGTCAAACGATAATTTAG‐3′.
Cryosection and Eosin Staining
Embryos were fixed in 4% PFA in PBS at 4°C for 16 h following X‐gal staining. After fixation, embryos were washed three times in PBS (5 min each, room temperature) and sequentially cryoprotected in 10%, 20%, and 30% sucrose in PBS. Samples were then embedded in O.C.T. compound (Tissue‐Tek O.C.T. compound, SAKURA Finetechnical Co. Ltd., 4583) and frozen for cryosectioning.
Cryosections were cut at a thickness of 12 μm using a cryostat (Leica Biosystems), mounted on glass slides, and air‐dried at 37°C for 30 min, followed by three washes in PBS (10 min each) to remove residual O.C.T. Sections were dehydrated through graded ethanol (70%, 90%, and 99%; 3 s each) and stained with eosin solution (1:5 dilution of pure eosin; Muto Pure Chemicals, No. 32042) in ethanol for 10 min. After brief dehydration in 99% ethanol (3 s), sections were mounted with Fluoromount/Plus (Diagnostic Biosystems, K048). Images were acquired using a light microscope (Keyence).
Author Contributions
Yukihiro Harada:methodology, data curation, investigation, validation, visualization, writing – review and editing, conceptualization.Teruhisa Kawamura:conceptualization, funding acquisition, writing – review and editing, project administration, supervision.Tomoe Ueyama:methodology, investigation, writing – review and editing.Osamu Nakagawa:conceptualization, funding acquisition, project administration, writing – review and editing.Yunce Wang:investigation, methodology, data curation, validation, visualization, writing – original draft, writing – review and editing.Daiki Seya:methodology, investigation.
Funding
This work was supported in part by the grants from Japan Society for the Promotion of Science (Grants‐in‐Aid for Scientific Research, Grant Numbers JP21H02890 (to O.N., T.K., and Y.H.), JP23K18268 (to O.N., T.K., and Y.H.), JP24K02430 (to O.N., T.K., and Y.H.), and JP24K22041 (to T.K., Y.H. and T.U.)).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Blackwood, E. M. , andJ. T. Kadonaga. 1998. “Going the Distance: A Current View of Enhancer Action. ”Science281, no. 5373: 60–63. . doi.org/10.1126/science.281.5373.60
- Charron, F. , andM. Nemer. 1999. “GATA Transcription Factors and Cardiac Development. ”Seminars in Cell & Developmental Biology10, no. 1: 85–91. . doi.org/10.1006/scdb.1998.0281
- Egbe, A. , S. Uppu, A. Stroustrup, S. Lee, D. Ho, andS. Srivastava. 2014. “Incidences and Sociodemographics of Specific Congenital Heart Diseases in The United States of America: An Evaluation of Hospital Discharge Diagnoses. ”Pediatric Cardiology35, no. 6: 975–982. . doi.org/10.1007/s00246-014-0884-8
- Feiner, L. , A. L. Webber, C. B. Brown, et al. 2001. “Targeted Disruption of Semaphorin 3C Leads to Persistent Truncus Arteriosus and Aortic Arch Interruption. ”Development128, no. 16: 3061–3070. . doi.org/10.1242/dev.128.16.3061
- Garry, D. J. , andE. N. Olson. 2006. “A Common Progenitor at the Heart of Development. ”Cell127, no. 6: 1101–1104. . doi.org/10.1016/j.cell.2006.11.031
- Heslop, J. A. , B. Pournasr, J. T. Liu, andS. A. Duncan. 2021. “GATA6 Defines Endoderm Fate by Controlling Chromatin Accessibility During Differentiation of Human‐Induced Pluripotent Stem Cells. ”Cell Reports35, no. 7: 109145. . doi.org/10.1016/j.celrep.2021.109145
- Hikspoors, J. P. J. M. , N. Kruepunga, G. M. C. Mommen, S. E. Köhler, R. H. Anderson, andW. H. Lamers. 2022. “A Pictorial Account of the Human Embryonic Heart Between 3. 5 and 8 Weeks of Development. ”Communications Biology5, no. 1: 1–22. . doi.org/10.1038/s42003-022-03153-x
- Ihara, D. , Y. Watanabe, D. Seya, et al. 2020. “Expression of Hey2 Transcription Factor in the Early Embryonic Ventricles Is Controlled Through a Distal Enhancer by Tbx20 and Gata Transcription Factors. ”Developmental Biology461, no. 2: 124–131. . doi.org/10.1016/j.ydbio.2020.02.001
- Kelly, R. G. 2023. “The Heart Field Transcriptional Landscape at Single‐Cell Resolution. ”Developmental Cell58, no. 4: 257–266. . doi.org/10.1016/j.devcel.2023.01.010
- Kelly, R. G. , M. E. Buckingham, andA. F. Moorman. 2014. “Heart Fields and Cardiac Morphogenesis. ”Cold Spring Harbor Perspectives in Medicine4, no. 10: 1–11. . doi.org/10.1101/cshperspect.a015750
- Kodo, K. , T. Nishizawa, M. Furutani, et al. 2009. “GATA6 Mutations Cause Human Cardiac Outflow Tract Defects by Disrupting Semaphorin‐Plexin Signaling. ”Proceedings of the National Academy of Sciences106, no. 33: 13933–13938. . doi.org/10.1073/pnas.0904744106
- Kodo, K. , S. Shibata, S. Miyagawa‐Tomita, et al. 2017. “Regulation of Sema3c and the Interaction Between Cardiac Neural Crest and Second Heart Field During Outflow Tract Development. ”Scientific Reports7, no. 1: 1–13. . doi.org/10.1038/s41598-017-06964-9
- Lepore, J. J. , P. A. Mericko, L. Cheng, et al. 2006. “GATA‐6 Regulates Semaphorin 3C and Is Required in Cardiac Neural Crest for Cardiovascular Morphogenesis. ”Journal of Clinical Investigation116, no. 4: 929–939. . doi.org/10.1172/JCI27363
- Li, L. , andZ. Wunderlich. 2017. “An Enhancer's Length and Composition Are Shaped by Its Regulatory Task. ”Frontiers in Genetics8, no. May: 63. . doi.org/10.3389/fgene.2017.00063
- Meilhac, S. M. , andM. E. Buckingham. 2018. “The Deployment of Cell Lineages That Form the Mammalian Heart. ”Nature Reviews Cardiology15, no. 11: 705–724. . doi.org/10.1038/s41569-018-0086-9
- Mitchell, S. C. , S. B. Korones, andH. W. Berendes. 1971. “Congenital Heart Disease in 56, 109 Births. Incidence and Natural History. ”Circulation43, no. 3: 323–332. . doi.org/10.1161/01.CIR.43.3.323
- Neeb, Z. , J. D. Lajiness, E. Bolanis, andS. J. Conway. 2013. “Cardiac Outflow Tract Anomalies. ”WIREs Developmental Biology2, no. 4: 499–530. . doi.org/10.1002/wdev.98
- Plein, A. , A. Calmont, A. Fantin, et al. 2015. “Neural Crest–Derived SEMA3C Activates Endothelial NRP1 for Cardiac Outflow Tract Septation. ”Journal of Clinical Investigation125, no. 7: 2661–2676. . doi.org/10.1172/JCI79668
- Rammah, M. , M. Théveniau‐Ruissy, R. Sturny, F. Rochais, andR. G. Kelly. 2022. “PPARγ and NOTCH Regulate Regional Identity in the Murine Cardiac Outflow Tract. ”Circulation Research131: 1–17. . doi.org/10.1161/CIRCRESAHA.122.320766
- Rana, M. S. , M. Théveniau‐Ruissy, C. De Bono, et al. 2014. “Tbx1 Coordinates Addition of Posterior Second Heart Field Progenitor Cells to the Arterial and Venous Poles of the Heart. ”Circulation Research115, no. 9: 790–799. . doi.org/10.1161/CIRCRESAHA.115.305020
- Reller, M. D. , M. J. Strickland, T. Riehle‐Colarusso, W. T. Mahle, andA. Correa. 2008. “Prevalence of Congenital Heart Defects in Metropolitan Atlanta, 1998–2005. ”Journal of Pediatrics153, no. 6: 807–813. . doi.org/10.1016/j.jpeds.2008.05.059
- Stefanovic, S. , B. Laforest, J. ‐P. Desvignes, et al. 2020. “Hox‐Dependent Coordination of Mouse Cardiac Progenitor Cell Patterning and Differentiation. ”eLife9: 1–32. . doi.org/10.7554/eLife.55124
- Théveniau‐Ruissy, M. , M. Dandonneau, K. Mesbah, et al. 2008. “The del22q11. 2 Candidate Gene Tbx1 Controls Regional Outflow Tract Identity and Coronary Artery Patterning. ”Circulation Research103, no. 2: 142–148. . doi.org/10.1161/CIRCRESAHA.108.172189
- Thom, T. , N. Haase, W. Rosamond, et al. 2006. “Heart Disease and Stroke Statistics—2006 Update. ”Circulation113, no. 6: e85–e151. . doi.org/10.1161/CIRCULATIONAHA.105.171600
- Valdembri, D. , D. Regano, F. Maione, E. Giraudo, andG. Serini. 2016. “Class 3 Semaphorins in Cardiovascular Development. ”Cell Adhesion & Migration10, no. 6: 641–651. . doi.org/10.1080/19336918.2016.1212805
- Van Der Linde, D. , E. E. M. Konings, M. A. Slager, et al. 2011. “Birth Prevalence of Congenital Heart Disease Worldwide: A Systematic Review and Meta‐Analysis. ”Journal of the American College of Cardiology58, no. 21: 2241–2247. . doi.org/10.1016/j.jacc.2011.08.025
- Verzi, M. P. , D. J. McCulley, S. De Val, E. Dodou, andB. L. Black. 2005. “The Right Ventricle, Outflow Tract, and Ventricular Septum Comprise a Restricted Expression Domain Within the Secondary/Anterior Heart Field. ”Developmental Biology287, no. 1: 134–145. . doi.org/10.1016/j.ydbio.2005.08.041
- Yamaguchi, N. , E. W. Chang, Z. Lin, et al. 2023. “An Anterior Second Heart Field Enhancer Regulates the Gene Regulatory Network of the Cardiac Outflow Tract. ”Circulation148, no. 21: 1705–1722. . doi.org/10.1161/CIRCULATIONAHA.123.065700
Republished from the open web under CC-BY. Authors: Wang Y, Harada Y, Ueyama T, Seya D, Nakagawa O, Kawamura T. Read the original.