Renal expression of Bach1 and oxidative stress in chronic kidney disease: evidence from a preclinical model.
Introduction The transcription factor BTB and CNC homology 1 (Bach1) represses the nuclear factor erythroid 2-related factor 2 (Nrf2), which controls antioxidant gene expression, and its role in chronic kidney disease (CKD) remains unclear. Methods CKD was induced by 5/6 nephrectomy in rats. Kidney fibrosis and oxidative stress markers were measured. The gene expression of Nrf2, Bach1, and nuclear factor kappa B (NF-κB) was assessed. Results CKD increased oxidative stress markers in plasma, kidney, and heart, as well as promoted kidney fibrosis. Moreover, CKD reduced cardiac Nrf2 expression. However, Bach1, Nrf2, and NF-κB remained unchanged in the kidney. Conclusion CKD did not modulate Bach1 mRNA levels in the kidneys of 5/6 nephrectomized rats.
INTRODUCTION
Reactive oxygen species (ROS) generation in chronic kidney disease (CKD) contributes to disease pathophysiology through direct renal cell injury, leading to functional impairment, inflammation, and fibrosis, as well as through the activation of signaling pathways and transcription factors that amplify these processes1. BTB and CNC homology 1 (Bach1) is a widely expressed transcription factor and a key regulator of the oxidative stress response2. Bach1 inhibits the nuclear factor erythroid 2–related factor 2 (Nrf2) by competing for the antioxidant response element, thereby suppressing antioxidant gene expression2,3and contributing to increased ROS, apoptosis, and cellular senescence4. While Nrf2 mediates cytoprotective responses, Bach1 acts as its primary negative regulator. Furthermore, the balance between oxidative stress and inflammation is regulated by crosstalk between the nuclear factor kappa B (NF-κB) and Nrf2 pathways, as Nrf2 activation can attenuate NF-κB–mediated inflammation, whereas NF-κB activation may suppress the antioxidant response5. Aging-associated increases in Bach1 impair redox homeostasis and exacerbate oxidative stress. In CKD, reduced Nrf2 expression and chronic inflammation are frequently observed. Thus, Bach1 inhibition may enhance Nrf2 activity and represent a potential therapeutic strategy2,3,4,6>.
Accordingly, this study evaluated oxidative stress, inflammatory markers, and renal structural alterations in a preclinical model of CKD.
METHODS
All procedures were conducted in accordance with the ARRIVE guidelines and complied with the criteria established by the National Council for the Control of Animal Experimentation (CONCEA) and were approved by the Animal Research Ethics Committee of theUniversidade Federal Fluminense(protocol number 956/2017).
Animals
Fourteen male Wistar rats were used and housed under controlled temperature (22 ± 1 °C) and humidity (60 ± 10%) conditions, with a 12:12 h light–dark cycle. Animals had free access to commercial chow and water and were weighed weekly. CKD was induced by 5/6 nephrectomy, as previously described7. The animals were randomly assigned to two groups: a control (Sham) group, which underwent surgical manipulation of the renal pedicles without nephrectomy (n = 7), and a CKD group subjected to 5/6 nephrectomy (n = 7).
Euthanasia
Animals were deeply anesthetized with ketamine (40 mg/kg) and xylazine (8 mg/kg) (Laboratório Virbac S.A., São Paulo, Brazil) and euthanized by cardiac puncture until total exsanguination. Kidney and heart tissues were carefully dissected and stored at -80 °C for further analysis.
Biochemical Analysis
Blood samples were drawn from each animal by cardiac puncture. Plasma and serum were separated by centrifugation (15 min, 4500 × g, 4 °C). Serum levels of creatinine and urea were measured using Bioclin®kits with the Bioclin BS-120 chemistry analyzer.
Lipid Peroxidation and Protein Carbonylation
Lipid peroxidation in renal and cardiac tissues was evaluated by the thiobarbituric acid reactive substances (TBARS) assay, and plasma protein oxidation was assessed by quantifying carbonyl groups using 2,4-dinitrophenylhydrazine (DNPH). Both analyses followed the methodology described by Costa et al.8. All samples were prepared in RIPA buffer containing a protease inhibitor cocktail (Sigma-Aldrich, Missouri, USA).
Kidney Morphology
Kidney sections were stained with Masson trichrome to evaluate tubulointerstitial fibrosis. Ten random fields of the tubulointerstitial area from five animals were selected and analyzed using the Image-Pro Plus (Media Cybernetics, Silver Spring, MD, USA) through the density threshold selection tool.
Gene Expression
Bach1,NF-κB,andNrf2mRNA expression was evaluated using real-time quantitative polymerase chain reaction (PCR), according to Leal et al.9. TaqMan Gene Expression Assays (Thermo Fisher Scientific) were used forBach1(Rn01477344_m1),NF-κB(Rn01399572_m1),Nrf2(Rn00477784_m1), and the control gene,Gapdh(Rn01775763_g1). Reactions were performed on the ABI Prism 7500 Sequence Detection System (Applied Biosystems) under standard conditions.Bach1expression was evaluated only in kidney tissue, andNF-κBandNrf2were evaluated in the kidney and heart.
Statistical Analysis
Results are presented as mean ± standard deviation, and the ROUT test was used to identify outliers. Data normality was assessed with the Shapiro–Wilk test. Group comparisons used an unpairedt-test, and a two-way ANOVA was applied for analyses over time. A multivariate approach was also employed. The variation in sample size for gene expression reflected limited biological material. A correlation matrix was generated, and principal component analysis (PCA) was conducted on standardized variables (z-score transformation) to account for differences in measurement scales and ensure equal contribution of all variables. Component retention followed the Kaiser–Guttman criterion, with eigenvalues ≥ 1.0 considered significant, and the variance explained by each retained component was reported. PCA constituted an exploratory, unsupervised dimensionality reduction method to identify dominant patterns and relationships among correlated variables, facilitating data interpretation and visualization rather than inferential testing. All analyses were performed using GraphPad Prism (version 10.2.3; GraphPad Software, Boston, MA, USA), and statistical significance was set atp< 0.05.
RESULTS
After 4 weeks, CKD was established. The CKD group showed significantly lower body mass than the control group (Figure 1A). As expected, the CKD group presented a significant increase in creatinine (1.06 ± 0.14 vs. 0.63 ± 0.05 mg/dL; p = 0.0006) and serum urea levels compared to the control group (60.29 ± 5.70 vs. 30.29 ± 2.98 mg/dL; p < 0.0001) (Figures 1Band1C). Additionally, plasma protein carbonyl levels were significantly elevated in the CKD group when compared to the control group (689.90 ± 42.06 vs. 584.80 ± 27.73 nmol/mg protein; p = 0.0003) (Figure 1D). Regarding lipid peroxidation, the CKD rats exhibited markedly higher levels in the kidney (0.27 ± 0.12 vs. 0.14 ± 0.02 μmol/mg tissue; p = 0.0451) and heart (2.46 ± 0.69 vs. 0.77 ± 0.40 μmol/mg tissue; p = 0.0055) compared to the control group (Figures 1Eand1F). Histological analysis of kidney photomicrographs revealed a marked increase in tubulointerstitial fibrosis in the CKD group relative to the control group (Figure 1G).
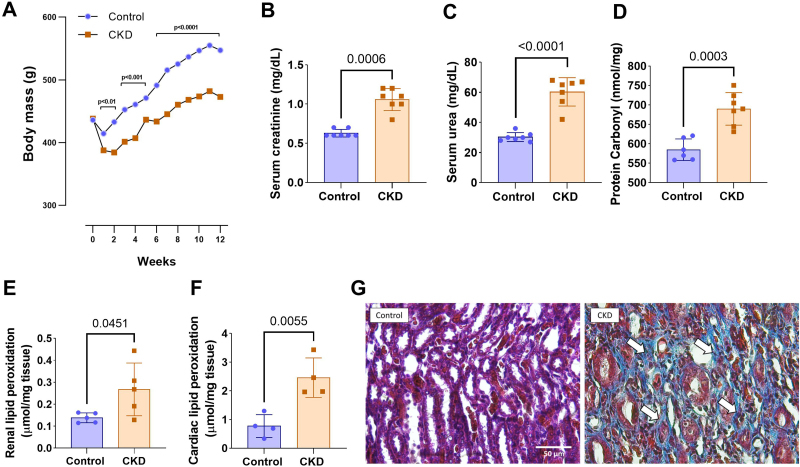
Body mass evolution over 12 weeks in control and CKD groups (A). Serum creatinine (B), serum urea (C), and protein carbonyl (D). Kidney (E) and heart (F) Lipid peroxidation levels in control and CKD animals. Representative photomicrographs of renal tissue from control and CKD animals (G). Histological analysis using Masson’s trichrome staining (×400 magnification) to evaluate tubulointerstitial fibrosis. Arrows (↑) indicate collagen deposition, markedly increased in the CKD group compared to the control group. (A), (F), and (G): control: n = 5; CKD: n = 5; (B) and (C): control: n = 7; CKD: n = 7; (D) control: n = 6; CKD: n = 7; (E): control: n = 4; CKD: n = 4.
There was no significant difference in renalBach1mRNA expression between groups (Figure 2A).NF-κBmRNA expression remained unchanged in both heart (p = 0.9982) and kidney (p = 0.7117) tissues (Figures 2Band2C). However, animals in the CKD group showed lower cardiacNrf2mRNA expression (0.71 ± 0.07 vs. 1.03 ± 0.24; p = 0.0241) compared to the control group (Figure 2D), whereas no difference was observed in the kidney (1.46 ± 1.53 vs. 0.92 ± 0.67; p = 0.3432) (Figure 2E).
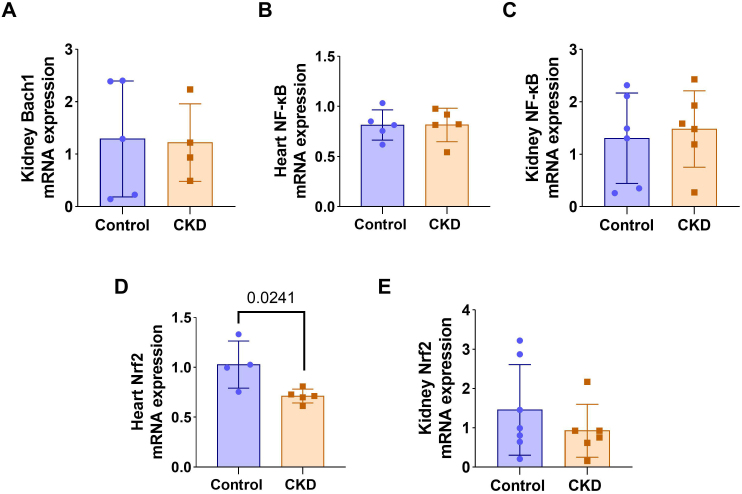
mRNA expression ofBach1in the kidney (A),Nf-κBin the heart (B),Nf-κBin the kidney (C),Nrf2in the heart (D), andNrf2in the kidney (E) in the experimental groups.
Multivariate analysis considered parameters related to inflammation, oxidative stress, and renal function. The heatmap matrix is presented inFigure 3A. PCA yielded two principal components, explaining 48.7% (PC1) and 28.0% (PC2) of the variance, accounting for a cumulative variance of 76.7% (Figure 3B). PC1 was strongly correlated with renalBach1,NF-κB, andNrf2expression, indicating an integrated response among these variables. PC2 was predominantly associated with renal lipid peroxidation levels and cardiacBach1,NF-κB, andNrf2expression (Figures 3Cand3D).
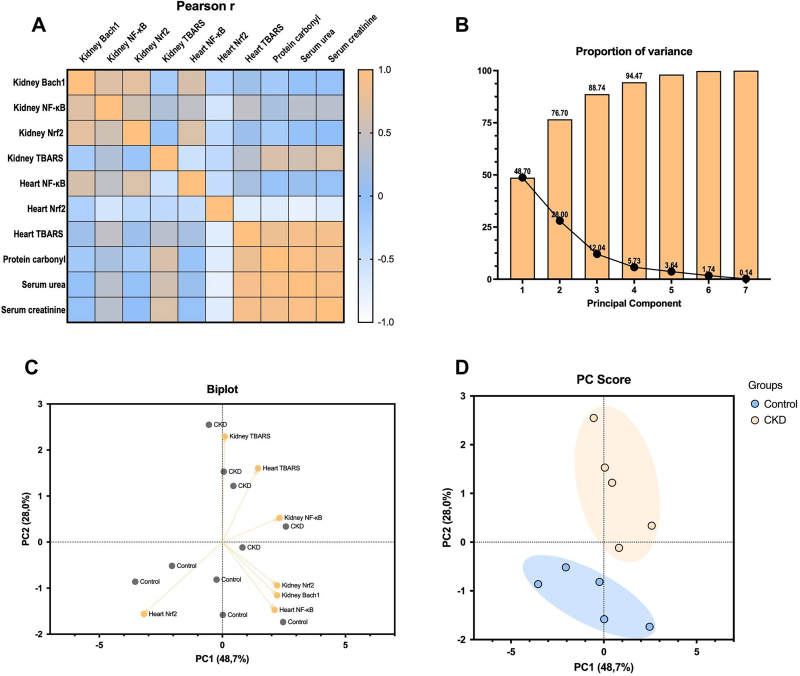
Principal component analysis. Pearson’s correlation heatmap (A). Cumulative variance of PCs (B). Biplot of loading scores (C). Score plots of PCs (D). Control: n = 5; CKD: n = 5.
DISCUSSION
The results of this study demonstrate that CKD in rats is associated with increased oxidative stress, as evidenced by elevated lipid peroxidation in renal and cardiac tissues and increased plasma protein carbonyl levels. In addition, CKD animals exhibited reduced Nrf2 mRNA expression in cardiac tissue, suggesting an impaired systemic antioxidant response. Notably,Bach1mRNA expression in the kidney did not differ significantly between groups, despite its established role as a negative regulator of Nrf2.
As activation of the Nrf2 pathway represents a therapeutic target in CKD, its downregulation has been associated with the progression of cardiometabolic disorders10,11. Nrf2 regulation involves complex molecular mechanisms influenced by multiple pathophysiological processes12. In the present study, Nrf2 mRNA levels differed between tissues, with reduced expression in the heart but not in the kidney, indicating tissue-specific regulation that may be associated with increased lipid peroxidation and protein carbonylation.
Despite the presence of CKD and associated oxidative stress and inflammation in our experimental model, no significant differences in renal Bach1 gene expression were observed between groups. This finding was unexpected, given that Bach1 is a redox-sensitive transcriptional repressor known to respond to oxidative stress by regulating antioxidant response elements, often acting in opposition to Nrf24,13. Previous studies have shown that, under acute oxidative conditions, such as rhabdomyolysis-induced acute kidney injury, Bach1 expression and subcellular localization can change dynamically, with nuclear export representing an early response to elevated heme levels14. In contrast, under chronic conditions such as CKD, Bach1 regulation may differ and be influenced by adaptive or compensatory mechanisms that help maintain stable expression levels despite sustained oxidative stress3,15. Thus, the absence of transcriptional changes in Bach1 does not negate its potential functional relevance, as its activity may be modulated by post-transcriptional and post-translational mechanisms, including changes in protein stability or subcellular localization16. Although the Bach1–Nrf2 interaction may be dynamically regulated over time, this study was limited to a single experimental time point; therefore, any temporal interpretation should therefore be made with caution. In addition, in the 5/6 nephrectomy model, increased tubulointerstitial fibrosis is observed, along with adaptive hyperfiltration and glomerular hypertrophy of residual nephrons to sustain excretory capacity17. These compensatory changes increase intraglomerular pressure and mechanical stress, promoting filtration barrier injury, oxidative stress, and activation of pro-inflammatory pathways18. Importantly, inflammatory activity may occur without changes in NF-κB gene expression, as NF-κB activation is largely regulated at the post-translational level19. Collectively, these mechanisms may have influenced renal Bach1 expression and impaired antioxidant responses, including Nrf2.
Finally, PCA analysis demonstrated that PC1 explained 48.7% of the total variance and represented the main axis of separation between groups. Sham animals clustered on the negative side, whereas the CKD group shifted toward positive values, capturing the dominant multivariate signature associated with CKD-related alterations. PC2 accounted for an additional 28.0% of the variance and reflected intragroup variability without compromising group separation, with both components explaining 76.7% of the total variance. Consistently, correlation analysis indicated that oxidative stress and inflammation were associated with renal dysfunction, as increased lipid and protein oxidation correlated with higher urea and creatinine levels. Furthermore,NF-κBexpression andNrf2downregulation were associated with a pro-inflammatory and antioxidant-deficient profile. The biplot further demonstrated a clear separation between the Sham and CKD groups, with the CKD group associated with oxidative stress and inflammatory markers and the Sham group associated with higher cardiacNrf2expression. Notably,Bach1expression did not correlate with the CKD group, indicating that this preclinical model is not associated with changes in its gene expression.
In conclusion, this study demonstrates that CKD promotes systemic oxidative stress and renal fibrosis, accompanied by reduced cardiac Nrf2 expression. Although renal Bach1 expression remained unchanged, future studies should explore its post-transcriptional regulation and the effects of Bach1 inhibition on Nrf2 signaling. Overall, these findings highlight the therapeutic potential of targeting redox-related pathways, particularly through Nrf2 activation, to mitigate CKD-associated tissue damage.
References
- KishiSNagasuHKidokoroKKashiharaNOxidative stress and the role of redox signalling in chronic kidney diseaseNat Rev Nephrol202420210119 doi.org/10.1038/s41581-023-00775-0
- ZhangXGuoJWeiXNiuCJiaMLiQBach1: function, regulation, and involvement in diseaseOxid Med Cell Longev2018201811347969 doi.org/10.1155/2018/1347969
- MafraDAlvarengaLCardozoLFMFStockler-PintoMBNakaoLSStenvinkelPInhibiting BTB domain and CNC homolog 1 (Bach1) as an alternative to increase Nrf2 activation in chronic diseasesBiochim Biophys Acta, Gen Subj202218666130129 doi.org/10.1016/j.bbagen.2022.130129
- MafraDAlvarengaLRibeiroMBaptistaBGFantonSPaivaBREvaluation of Bach1 mRNA expression in patients with chronic kidney disease: A preliminary studyHemodial Int20232733017 doi.org/10.1111/hdi.13084
- CasperEThe crosstalk between Nrf2 and NF-κB pathways in coronary artery disease: can it be regulated by SIRT6?Life Sci2023330122007 doi.org/10.1016/j.lfs.2023.122007
- YuCXiaoJHThe Keap1-Nrf2 system: a mediator between oxidative stress and agingOxid Med Cell Longev2021202116635460 doi.org/10.1155/2021/6635460
- AlmeidaPPThomasiBBMMenezesÁCCruzBOCostaNSBritoML5/6 nephrectomy affects enteric glial cells and promotes impaired antioxidant defense in the colonic neuromuscular layerLife Sci2022298120494 doi.org/10.1016/j.lfs.2022.120494
- CostaNSAraujoJRMeloMFSMotaJCAlmeidaPPCoutinho-WolinoKSEffects of probiotic-enriched minas cheese (Lactobacillus acidophilus La-05) on cardiovascular parameters in 5/6 nephrectomized ratsProbiotics Antimicrob Proteins202517287387 doi.org/10.1007/s12602-023-10173-4
- LealVOSaldanhaJFStockler-PintoMBCardozoLFMFSantosFRAlbuquerqueASDNRF2 and NF-κB mRNA expression in chronic kidney disease: a focus on nondialysis patientsInt Urol Nephrol20154712198591 doi.org/10.1007/s11255-015-1135-5
- Juul-NielsenCShenJStenvinkelPScholzeASystematic review of the nuclear factor erythroid 2-related factor 2 (NRF2) system in human chronic kidney disease: Alterations, interventions and relation to morbidityNephrol Dial Transplant202237590416 doi.org/10.1093/ndt/gfab031
- ArefinSMudrovcicNHobsonSPietrocolaFEbertTWardLJEarly vascular aging in chronic kidney disease: focus on microvascular maintenance, senescence signature and potential therapeuticsTransl Res20252753247 doi.org/10.1016/j.trsl.2024.11.001
- Guerrero-HueMRayego-MateosSVázquez-CarballoCPalomino-AntolínAGarcía-CaballeroCOpazo-RiosLProtective role of Nrf2 in renal diseaseAntioxidants202010139 doi.org/10.3390/antiox10010039
- YusoffFMMaruhashiTKawanoKINakashimaAChayamaKTashiroSBach1 plays an important role in angiogenesis through regulation of oxidative stressMicrovasc Res2021134104126 doi.org/10.1016/j.mvr.2020.104126
- YamaokaMShimizuHTakahashiTOmoriEMorimatsuHDynamic changes in Bach1 expression in the kidney of rhabdomyolysis-associated acute kidney injuryPLoS One2017127e0180934 doi.org/10.1371/journal.pone.0180934
- WeiXHeYYuYTangSLiuRGuoJThe multifaceted roles of BACH1 in disease: implications for biological functions and therapeutic applicationsAdv Sci20251210e2412850 doi.org/10.1002/advs.202412850
- IgarashiKWatanabe-MatsuiMWearing red for signaling: the heme-bach axis in heme metabolism, oxidative stress response and iron immunologyTohoku J Exp Med2014232422953 doi.org/10.1620/tjem.232.229
- NogueiraAPiresMJOliveiraPAPathophysiological mechanisms of renal fibrosis: a review of animal models and therapeutic strategiesIn Vivo2017311122 doi.org/10.21873/invivo.11019
- DwivediSSikarwarMSDiabetic nephropathy: pathogenesis, mechanisms, and therapeutic strategiesHorm Metab Res2025571717 doi.org/10.1055/a-2435-8264
- WonMByunHSParkKAHurGMPost-translational control of NF-κB signaling by ubiquitinationArch Pharm Res2016398107584 doi.org/10.1007/s12272-016-0772-2
Republished from the open web under CC-BY. Authors: Cruz BOD, Almeida PP, Silva-Costa N, Brito ML, Magliano DC, Stockler-Pinto MB. Read the original.